CELL AND ITS COMPONENTS

Cell and Its Components
Cells are the body’s smallest structural and functional units. They are grouped together to form tissues, each of which has a specialized function, e.g. blood, muscle, bone. Different tissues are grouped together to form organs, e.g. the heart, stomach and brain.
Organs are grouped together to form systems, each of which performs a particular function that maintains homeostasis and contributes to the health of the individual.
A cell consists of a plasma membrane enclosing a number of organelles suspended in a watery fluid called cytosol.

1. CYTOSOL:
The cytosol (intracellular fluid) is the fluid portion of the cytoplasm that surrounds organelles and constitutes about 55% of total cell volume.
Although it varies in composition and consistency from one part of a cell to another, cytosol is 75–90% water plus various dissolved and suspended components.
Among these are different types of ions, glucose, amino acids, fatty acids, proteins, lipids, ATP, and waste products.
The cytosol is the site of many chemical reactions required for a cell’s existence
2. ORGANELLES:
Organelles are specialized structures within the cell that have characteristic shapes; they perform specific functions in cellular growth, maintenance, and reproduction.
They include: the nucleus, mitochondria, ribosomes, endoplasmic reticulum, Golgi apparatus, lysosomes and the cytoskeleton. The cell content excluding the nucleus is the cytoplasm, i.e. the cytosol and other organelles.
3. THE CYTOSKELETON:
The cytoskeleton is a network of protein filaments that extends throughout the cytosol. Three types of filamentous proteins contribute to the cytoskeleton’s structure, as well as the structure of other organelles.
Functions:
1. Serves as a scaffold that helps to determine a cell’s shape and to organize the cellular contents.
2. Aids movement of organelles within the cell, of chromosomes during cell division, and of whole cells such as phagocytes.
In the order of their increasing diameter, these structures are:
Microfilaments:
These are the thinnest elements of the cytoskeleton. They are composed of the protein actin, and are most prevalent at the edge of a cell. Microfilaments have two general functions:
They help generate movement and provide mechanical support that is responsible for the basic strength and shapes of cells.

Intermediate filaments:
these filaments are thicker than microfilaments but thinner than microtubules.
Several different proteins can compose intermediate filaments, which are exceptionally strong.
They are found in parts of cells subject to mechanical stress, help stabilize the position of organelles such as the nucleus, and help attach cells to one another.

Microtubules:
These are the largest of the cytoskeletal components and are long, unbranched hollow tubes composed mainly of the protein tubulin.
The assembly of microtubules begins in an organelle called the centrosome. Microtubules help determine cell shape.
They also function in the movement of organelles such as secretory vesicles, of chromosomes during cell division, and of specialized cell projections, such as cilia and flagella.

4. CENTROSOME:
The centrosome, located near the nucleus, consists of two components: a pair of centrioles and pericentriolar
material.
The two centrioles are cylindrical structures, each composed of nine clusters of three microtubules (triplets) arranged in a circular pattern.
The long axis of one centriole is at a right angle to the long axis of the other.
Surrounding the centrioles is pericentriolar material, which contains hundreds of ring-shaped complexes composed of the protein tubulin.
These tubulin complexes are the organizing centers for growth of the mitotic spindle, which plays a critical role in cell division, and for microtubule formation in nondividing cells.
During cell division, centrosomes replicate so that succeeding generations of cells have the capacity for cell division.
Function: The pericentriolar material of the centrosome contains tubulins that build microtubules in nondividing cells and form the mitotic spindle during cell division.

5. CILIA AND FLAGELLA:
Microtubules are the dominant components of cilia and flagella, which are motile projections of the cell surface.
Cilia are numerous, short, hair like projections that extend from the surface of the cell. Each cilium contains a core of 20 microtubules surrounded by plasma membrane.
Flagella are similar in structure to cilia but are typically much longer. Flagella usually move an entire cell.
A flagellum generates forward motion along its axis by rapidly wiggling in a wavelike pattern.
The only example of a flagellum in the human body is a sperm cell’s tail, which propels the sperm toward the oocyte in the uterine tube.
Functions:
1. Cilia move fluids along a cell’s surface.
2. A flagellum moves an entire cell.
6. RIBOSOMES:
Ribosomes are the sites of protein synthesis. The name of these tiny organelles reflects their high content of one type of ribonucleic acid.
Structurally, a ribosome consists of two subunits, one about half the size of the other.
The large and small subunits are made separately in the nucleolus, a spherical body inside the nucleus.
Once produced, the large and small subunits exit the nucleus separately, then come together in the cytoplasm.
Some ribosomes are attached to the outer surface of the nuclear membrane and to an extensively folded membrane called the endoplasmic reticulum.
These ribosomes synthesize proteins destined for specific organelles, for insertion in the plasma membrane, or for export from the cell.
Other ribosomes are “free” or unattached to other cytoplasmic structures.
Free ribosomes synthesize proteins used in the cytosol.
Ribosomes are also located within mitochondria, where they synthesize mitochondrial proteins.
Functions:
1. Ribosomes associated with endoplasmic reticulum synthesize proteins destined for insertion in the plasma membrane or secretion from the cell.
2. Free ribosomes synthesize proteins used in the cytosol.

7. ENDOPLASMIC RETICULUM:
The endoplasmic reticulum or ER is a network of membranes in the form of flattened sacs or tubules. The
ER extends from the nuclear envelope (membrane around the nucleus), to which it is connected, throughout the cytoplasm.
Cells contain two distinct forms of ER, which differ in structure and function. Rough ER is continuous with the nuclear membrane and usually is folded into a series of flattened sacs.
The outer surface of rough ER is studded with ribosomes, the sites of protein synthesis.
Rough ER produces secretory proteins, membrane proteins, and many organellar proteins.

Smooth ER extends from the rough ER to form a network of membrane tubules. Unlike rough ER, smooth ER does not have ribosomes on the outer surfaces of its membrane.
Smooth ER does not synthesize proteins, but it does synthesize fatty acids and steroids, such as estrogens and testosterone.
In liver cells, enzymes of the smooth ER help release glucose into the bloodstream and inactivate or detoxify lipid-soluble drugs or potentially harmful substances.

Functions:
1. Rough ER synthesizes glycoproteins and phospholipids that are transferred into cellular organelles, inserted into the plasma membrane, or secreted during exocytosis.
2. Smooth ER synthesizes fatty acids and steroids, such as estrogens and testosterone; inactivates or detoxifies drugs and other potentially harmful substances; removes the phosphate group from glucose-6phosphate; and stores and releases calcium ions that trigger contraction in muscle cells.
8. GOLGI COMPLEX:
Most of the proteins synthesized by ribosomes attached to rough ER are ultimately transported to other regions of the cell.
The first step in the transport pathway is through an organelle called the Golgi complex. It consists of 3 to 20 cisternae, small, flattened membranous sacs with bulging edges that resemble a stack of pita bread.
The cisternae are often curved, giving the Golgi complex a cuplike shape.

The convex entry or cis face is a cisterna that faces the rough ER. The concave exit or trans face is a cisterna that faces the plasma membrane. Sacs between the entry and exit faces are called medial cisternae.
Functions:
1. Modifies, sorts, packages, and transports proteins received from the rough ER.
2. Forms secretory vesicles that discharge processed proteins via exocytosis into extracellular fluid; forms membrane vesicles that ferry new molecules to the plasma membrane; forms transport vesicles that carry molecules to other organelles, such as lysosomes.
8. LYSOSOMES:
Lysosomes are membrane-enclosed vesicles that form from the Golgi complex.
A lysosome can engulf another organelle, digest it, and return the digested components to the cytosol for reuse. In this way, old organelles are continually replaced.
The process by which entire worn-out organelles are digested is called autophagy.

Functions:
1. Digest substances that enter a cell via endocytosis and transport final products of digestion into cytosol.
2. Carry out autophagy, the digestion of worn-out organelles.
3. Carry out autolysis, the digestion of entire cell.
4. Carry out extracellular digestion.
9. PEROXISOMES:
Another group of organelles similar in structure to lysosomes, but smaller, are the peroxisomes
Peroxisomes, also called microbodies, contain several oxidases, enzymes that can oxidize (remove hydrogen atoms from) various organic substances.

10. PROTEASOMES:
Lysosomes degrade proteins delivered to them in vesicles.
Cytosolic proteins also require disposal at certain times in the life of a cell.
Continuous destruction of unneeded, damaged, or faulty proteins is the function of tiny barrel-shaped structures consisting of four stacked rings of proteins around a central core called proteasomes.

Proteasomes were so named because they contain myriad proteases, enzymes that cut proteins into small peptides.
11. MITOCHONDRIA:
Mitochondria are referred to as the “powerhouses” of the cell.
A mitochondrion consists of an outer mitochondrial membrane and an inner mitochondrial membrane with a small fluid-filled space between them.
Both membranes are similar in structure to the plasma membrane.
The inner mitochondrial membrane contains a series of folds called cristae. The central fluidfilled cavity of a mitochondrion, enclosed by the inner mitochondrial membrane, is the matrix.
The elaborate folds of the cristae provide an enormous surface area for the chemical reactions that are part of the aerobic phase of cellular respiration, the reactions that produce most of a cell’s ATP.
The enzymes that catalyze these reactions are located on the cristae and in the matrix of the mitochondria.

12. NUCLEUS:
The nucleus is a spherical or oval-shaped structure that usually is the most prominent feature of a cell.
A double membrane called the nuclear envelope separates the nucleus from the cytoplasm. Both layers of the nuclear envelope are lipid bilayers similar to the plasma membrane.
The outer membrane of the nuclear envelope is continuous with rough ER and resembles it in structure.
Many openings called nuclear pores extend through the nuclear envelope. Each nuclear pore consists of a circular arrangement of proteins surrounding a large central opening that is about 10 times wider than the pore of a channel protein in the plasma membrane. Nuclear pores control the movement of substances between the nucleus and the cytoplasm.
Inside the nucleus are one or more spherical bodies called nucleoli that function in producing ribosomes. Each nucleolus is simply a cluster of protein, DNA, and RNA; it is not enclosed by a membrane.
Nucleoli are the sites of synthesis of rRNA and assembly of rRNA and proteins into ribosomal subunits.
Nucleoli are quite prominent in cells that synthesize large amounts of protein, such as muscle and liver cells. Nucleoli disperse and disappear during cell division and reorganize once new cells are formed.
Within the nucleus are most of the cell’s hereditary units, called genes, which control cellular structure and direct cellular activities.
Genes are arranged along chromosomes.
Human somatic (body) cells have 46 chromosomes, 23 inherited from each parent.
Each chromosome is a long molecule of DNA that is coiled together with several proteins. This complex of DNA, proteins, and some RNA is called chromatin. The total genetic information carried in a cell or an organism is its genome.

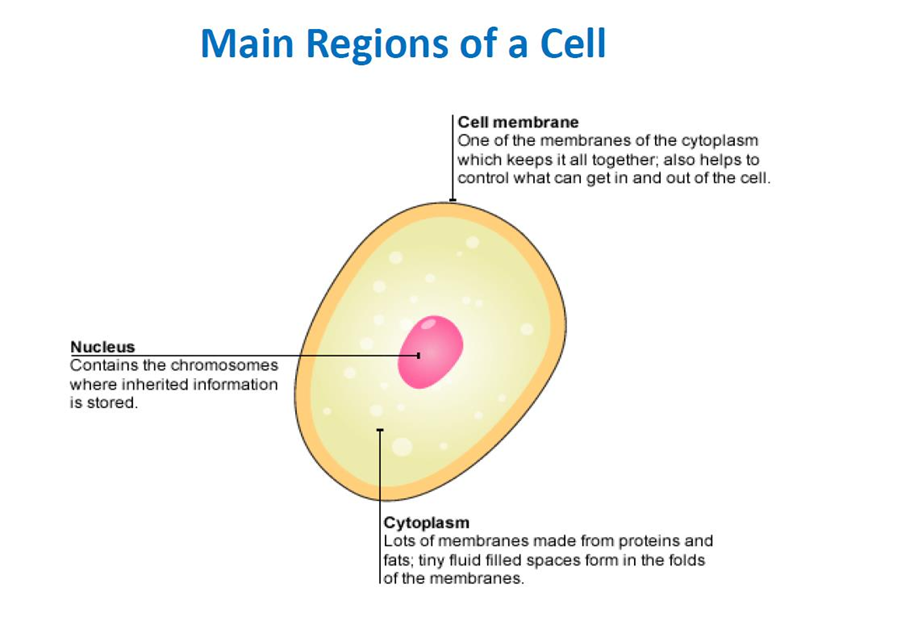
13. PLASMA MEMBRANE:
The plasma membrane, a flexible yet sturdy barrier that surrounds and contains the cytoplasm of a cell, is described by using a structural model called the fluid mosaic model.

Structure of the Plasma Membrane:
The Lipid Bilayer: The basic structural framework of the plasma membrane is the lipid bilayer, two back-to-back layers made up of three types of lipid molecules—phospholipids, cholesterol, and glycolipids. About 75% of the membrane lipids are phospholipids (lipids that contain phosphorus).
Present in smaller amounts are cholesterol (about 20%), a steroid with an attached -OH (hydroxyl) group, and various glycolipids (about 5%), lipids with attached carbohydrate groups.
The bilayer arrangement occurs because the lipids are amphipathic molecules, which means that they have both polar and nonpolar parts.
In phospholipids, the polar part is the phosphate containing “head,” which is hydrophilic (water loving). The nonpolar parts are the two long fatty acid “tails,” which are hydrophobic (water fearing) hydrocarbon chains. Because “like seeks like,” the phospholipid molecules orient themselves in the bilayer with their hydrophilic heads facing outward.
In this way, the heads face a watery fluid on either side—cytosol on the inside and extracellular fluid on the outside. The hydrophobic fatty acid tails in each half of the bilayer point toward one another, forming a nonpolar, hydrophobic region in the membrane’s interior.

Arrangement of Membrane Proteins:
Membrane proteins are classified as integral or peripheral according to whether they are firmly embedded in the membrane.
Integral proteins
extend into or through the lipid bilayer among the fatty acid tails and are firmly embedded in it. Most integral proteins are transmembrane proteins, which means that they span the entire lipid bilayer and protrude into both the cytosol and extracellular fluid.
Peripheral proteins
are not as firmly embedded in the membrane. They associate more loosely with the polar heads of membrane lipids or with integral proteins at the inner or outer surface of the membrane.
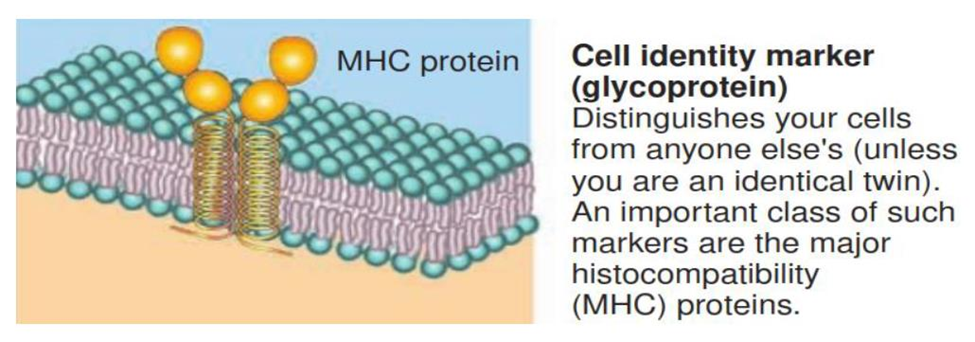
Many membrane proteins are glycoproteins, proteins with carbohydrate groups attached to the ends that protrude into the extracellular fluid.
The carbohydrate portions of glycolipids and glycoproteins form an extensive sugary coat called the glycocalyx.
The pattern of carbohydrates in the glycocalyx, varies from one cell to another.
Therefore, the glycocalyx acts like a molecular “signature” that enables cells to recognize one another.
Functions of Membrane Proteins:
Some integral membrane proteins form ion channels, pores or holes through which specific ions, such as potassium ions, can flow to get into or out of the cell.

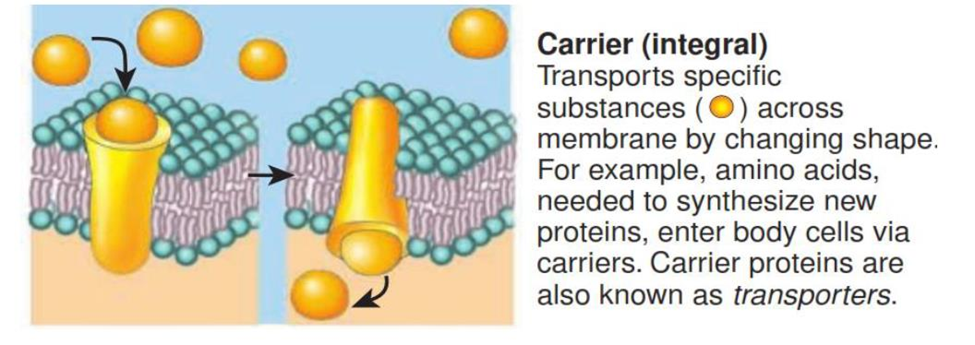
Other integral proteins act as carriers, selectively moving a polar substance or ion from one side of the membrane to the other. Carriers are also known as transporters.
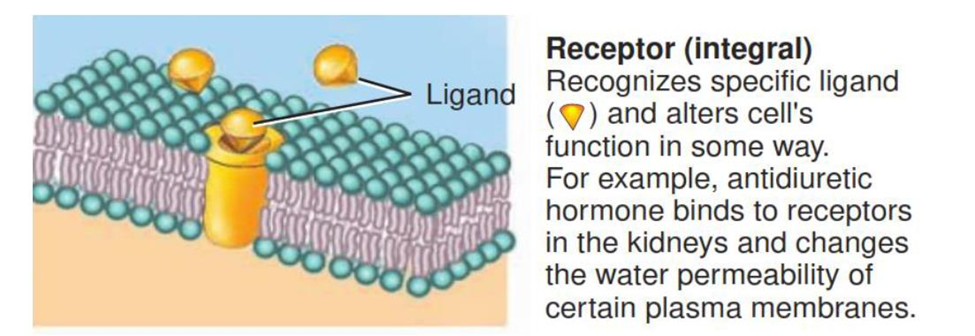
Integral proteins called receptors serve as cellular recognition sites. Each type of receptor recognizes and binds a specific type of molecule. For instance, insulin receptors bind the hormone insulin. A specific molecule that binds to a receptor is called a ligand of that receptor.
Some integral proteins are enzymes that catalyze specific chemical reactions at the inside or outside surface of the cell.

Integral proteins may also serve as linkers, which anchor proteins in the plasma membranes of neighboring cells to one another or to protein filaments inside and outside the cell. Peripheral proteins also serve as enzymes and linkers.

Membrane glycoproteins and glycolipids often serve as cell identity markers. They may enable a cell to recognize other cells of the same kind during tissue formation or to recognize and respond to potentially dangerous foreign cells. The ABO blood type markers are one example of cell identity markers.
Peripheral proteins help support the plasma membrane, anchor integral proteins, and participate in mechanical activities such as moving materials and organelles within cells, changing cell shape in dividing and muscle cells, and attaching cells to one another.
Also, Visit:
B. Pharma Notes | B. Pharma Notes | Study material Bachelor of Pharmacy pdf